Reviving,and,characterizing,three,species,of,dinof,lagellate,cysts,dormant,for,about,70,years,in,the,East,China,Sea:Biecheleria,brevisulcata,,Biecheleriopsis,adriatica,,and,Scrippsiella,donghaienis*
Zhangxi HU , Xiaoying SONG , Jinxiu WANG , Zhe TAO , Yuanyuan SUN ,Yuhang LI , Yuyang LIU , Yunyan DENG ,3,6, Lixia SHANG ,3,6, Zhaoyang CHAI ,3,6,Yingzhong TANG ,3,6,**
1 College of Fisheries, Guangdong Ocean University, Zhanjiang 524088, China
2 CAS Key Laboratory of Marine Ecology and Environmental Sciences, Institute of Oceanology, Chinese Academy of Sciences,Qingdao 266071, China
3 Laboratory of Marine Ecology and Environmental Science, Pilot National Laboratory for Marine Science and Technology(Qingdao), Qingdao 266237, China
4 CAS Key Laboratory of Experimental Marine Biology, Institute of Oceanology, Chinese Academy of Sciences, Qingdao 266071,China
5 Laboratory of Marine Organism Taxonomy and Phylogeny, Institute of Oceanology, Chinese Academy of Sciences, Qingdao 266071, China
6 Center for Ocean Mega-Science, Chinese Academy of Sciences, Qingdao 266071, China
7 University of Chinese Academy of Sciences, Beijing 100049, China
Abstract Many marine dinof lagellates can form resting cysts as a part of their life cycle, and the cysts could be buried in sediment and remained viable for as long as over 150 years. However, only a very limited number of cyst species have been revived from long-buried sediments and investigated in regard to a possible shift in the intra-specif ic genetic structure of a species detected from the historical record at a particular location. Here, we report a successful germination of three species of resting cysts that were sampled from the depth dated back to 1941±18 AD from a 44-cm sediment core from the East China Sea.Seven isolates were established from germination of single cyst isolation or multi-cyst germinations. LSU rRNA gene or ITS sequences of these strains were obtained, then they were identif ied to be Biecheleria brevisulcata (f ive strains), Biecheleriopsis adriatica (one strain), and Scrippsiella donghaienis (one strain)in terms of morphology and rRNA gene sequence. Biecheleria brevisulcata strain 1, Bps. adriatica strain 21, and S. donghaienis strain 23 were examined in detail with light microscope (LM) and scanning electron microscope (SEM), and analyzed with high performance liquid chromatography (HPLC) for their pigment compositions, and genetic diversity. We also conf irmed the presence of a resting cyst of Bps. adriatica in the f ield for the f irst time. The LSU rRNA gene-based genetic distances of Bps. adriatica from that obtained from water sample, single-cell PCR sequencing for the cysts isolated from the surface sediment of the same sea area and that reported from other regions during the recent years, and ITS-based genetic distances of S. donghaienis from that obtained from cysts isolated from the surface sediment of the same location and that reported from other regions during the recent years indicated that the intra-specif ic genetic structure of each species in the sampling area may have shifted during the last 70 years. Our work conf irms that B.brevisulcata, Bps. adriatica, and S. donghaienis, all described as new species around 2010, have inhabited the East China Sea for about 70 years. The present work reports for the f irst time the revival of dinof lagellate resting cysts long-buried in the coastal sediments of China, which facilitates further study on the historical occurrences of other harmful dinof lagellates and their relevance to the regional climate and environmental changes in China.
Keyword: core sediment; dinof lagellate resting cyst; germination; Biecheleria brevisulcata; Biecheleriopsis adriatica; Scrippsiella donghaienis
Dinof lagellates constitute one of the main groups of marine phytoplankton in terms of their important contribution to the primary production, and comprise about 2 400 species belonging to 259 genera (Gómez,2012), which is still growing as more new taxa are to be described (Gómez et al., 2015; Takahashi et al.,2015, 2017, 2019; Boutrup et al., 2017; Luo et al.,2018; Hu et al., 2020b, 2021; Ok et al., 2021; Gu et al., 2022). The appearance of dinof lagellates dates back more than 400 million years in the fossil record,and this group of protists has evolved diverse features to adapt to their environments, including diverse morphologies, multiple-membrane cell walls, diff erent modes of nutrition, pigments, toxins, asexual and sexual reproduction, and other characters (Steidinger and Meave del Castillo, 2018). Among these features,the ability of about 10% of dinof lagellate species to produce resting cysts as a part of their life cycle is a vital one (Head, 1996; Bravo and Figueroa, 2014;Tang et al., 2016, 2021). It is now widely accepted that resting cysts play important roles in the biology and ecology of dinof lagellates (Anderson and Wall,1978; Dale, 2001; Bravo and Figueroa, 2014; Tang et al., 2016, 2021; Ellegaard and Ribeiro, 2018;Figueroa et al., 2018).
In the past, f ield studies on dinof lagellate cysts are generally focused on the surface sediments for mapping the distribution and abundance of cysts of important species, investigating species diversity of cyst assemblage in a region of concern, and conf irming the cyst presence for some species with particular importance (Luo et al., 2018; Limoges et al., 2020; Liu et al., 2020a, b, 2021; Mertens et al.,2020; Van Nieuwenhove et al., 2020; Hu et al., 2021,2022), whereas sediment cores representing a valuable archive of phytoplankton communities are usually used to reconstruct past environmental changes(Keafer et al., 1992; Dai et al., 2012; Ellegaard et al.,2013, 2020; Bringué et al., 2016; García-Moreiras et al., 2018; Kim et al., 2018; Price et al., 2018; de Freitas et al., 2020; Li et al., 2021; Siano et al., 2021).Cultures established from reviving dinof lagellate resting cysts collected from sediment cores have been used to evaluate the impact of environmental changes on the physiology, genetic structure, and diversity in various species (Ribeiro et al., 2011; Klouch et al.,2016; Lundholm et al., 2017; Kremp et al., 2018;Delebecq et al., 2020; Ellegaard et al., 2020; Girault et al., 2021). However, the numbers of dinof lagellate cysts remaining viable in long-buried sediments and the mechanisms behind it still need to be explored.Therefore, mining cyst records of dinof lagellate species buried in sediment cores is important in many aspects of the ecology of dinof lagellates and in reconstruction of the history of marine environmental changes.
Recently, we successfully established seven clonal cultures ofBiecheleriabrevisulcata,Biecheleriopsisadriatica, andScrippsielladonghaienisvia cyst germination from the depth dated back to 1941±18 AD of a sediment core collected from the East China Sea, and further characterized their morphologies,pigment compositions, and the genetic diversity in their LSU rRNA gene and ITS sequences.
2.1 Sediment core and surface sediment sampling and dating
One sediment core (S06-2, 120.417°E, 26.122°N)was collected from the East China Sea in September 19, 2018 during the public cruise of R/VXiangYangHong18organized by the National Natural Science Foundation of China and the First Institute of Oceanography, Ministry of Natural Resources,China, and one surface sediment sample (0–2 cm;S01-1, 122.997°E, 31.000°N) was also collected from the East China Sea in September 17, 2019 during the public cruise of R/VXiangYangHong18.The sediment core was sliced into 2-cm layers (the top 20 cm) and 4-cm layers (20 cm to the bottom)by caution. A total of 16 subsamples were collected for the210Pb and137Cs measurements. The detailed measurement and age determination was reported in Liu et al. (2021), and the standard error of age was produced in regression uncertainties.
2.2 Culture establishment
Cyst assemblage in the subsample (42–44 cm) of core sediment (S06-2) and surface sediment sample(S01-1) was concentrated using sodium polytungstate solution (SPT) (Bolch, 1997). For subsample of core sediment, single cysts were washed at least three times using sterile seawater (with a salinity of 31)enriched with f/2-Si medium (Guillard, 1975), and then micropipetted to a 24-well culture plate with each well containing 2.5-mL fresh medium and 2%antibiotic solution (a mixture of 10 000-IU penicillin and 10 000-μg/mL streptomycin; Solarbio, Beijing,China). The rest of cyst assemblage was transferred to a 6-well culture plate with each well containing 10-mL fresh medium and 2% antibiotic solution. The plates were incubated at 21 °C, 12-h꞉12-h light꞉dark cycle, and ~100 μmol photons/(m2·s). Cysts and new germlings were observed every day or every other day with an inverted microscope (IX73, Olympus,Japan) and photographed by a DP80 digital camera(Olympus, Japan). Five strains ofB.brevisulcata(S1,S2, S3, S4, and S5) were established from the cyst assemblage germination experiment, but the resting and empty cysts ofB.brevisulcatawere not observed.One strain ofBps.adriatica(S21) and one strain ofS.donghaienis(S23) were established from single cysts germination experiments, their resting and empty cysts were clearly recorded using an inverted microscope (IX73, Olympus, Japan) equipped with a DP80 digital camera (Olympus, Japan). All cultures were routinely maintained in the same condition mentioned above.
2.3 Light microscopic observation
Live cells ofB.brevisulcata(strain S1),Bps.adriatica(strain S21), andS.donghaienis(strain S23)were observed and photographed using a Zeiss Imager Z2 (Carl Zeiss, Gottingen, Germany) equipped with diff erential interference contrast (DIC), or an inverted microscope (IX73, Olympus, Japan) equipped with a digital camera (DP80, Olympus, Japan). For observation of thecal plates ofS.donghaienis, live cells were stained with Calcof luor White (Sigma-Aldrich, St. Louis, MO, USA) and examined using an epif luorescence microscope (BX53, Olympus,Japan) with a UV f ilter set (Fritz and Triemer,1985). Cells sizes ofB.brevisulcata,Bps.adriatica,andS.donghaienisfor 50 live cells at the midexponential growth phase were measured at ×400 (forB.brevisulcataandBps.adriatica), and ×200 (forS.donghaienis) magnif ication using a DP80 digital camera (Olympus, Tokyo, Japan).
2.4 SEM observation
For SEM observation, vegetative cells ofB.brevisulcataandBps.adriaticaat mid-exponential growth stage were f ixed with OsO4(2% f inal concentration), andS.donghaieniswith glutaraldehyde(2.5% f inal concentration) for 40–50 min. Fixed cells were gently f iltered onto 5-μm (B.brevisulcataandBps.adriatica) and 11-μm (S.donghaienis) pore size Millipore nylon membranes, dehydrated in an acetone series (10%, 30%, 50%, 70%, 90%, and three times in 100%, 15 min for each step), and critical pointdried with liquid CO2(EM CPD300, Leica, Austria).They were sputter-coated with platinum-palladium(EM ACE200, Leica, Austria), and observed using an S-3400N SEM (Hitachi, Japan).
2.5 Pigment analyses
Fifty milliliter of each culture ofB.brevisulcata(strain S1),Bps.adriatica(strain S21), andS.donghaienis(strain S23) in exponential growth(in cell densities of ca. 102 150, 95 900, and 980 cells/mL, respectively) were f iltered through 25-mm diameter glass f iber f ilter (Whatman, Maidstone,UK) and immediately frozen at -80 °C for later analyses. Pigments were analyzed on an Alliance HPLC (e2695, Waters, Milford, Massachusetts,USA) using a 100-μL sample injection according to Kong et al. (2012) and Hu et al. (2020a). Pigments were identif ied and quantif ied using Shimadzu Class-VP software and by comparing pigment spectra and retention times with those of 26 standard pigments(DHI Water and Environment, Hørsholm, Denmark;Kong et al., 2012; Hu et al., 2020a).
2.6 DNA extraction, PCR amplif ication, and rDNA-based phylogenetic analyses
Genomic DNA ofB.brevisulcata,Bps.adriatica,andS.donghaieniswere extracted using a plant DNA extraction kit (Tiangen, Beijing, China) according to the manufacturer’s protocol. ForB.brevisulcata,Bps.adriatica, andS.donghaienis, about 1 400 bp of LSU rDNA were amplif ied using primers of D1R(forward, 5ʹ-ACCCGCTGAATTTAAGCATA-3ʹ)(Scholin et al., 1994) and 28-1483R (reverse,5ʹ-GCTACTACCACCAAGATCTGC-3ʹ) (Daugbjerg et al., 2000), and forS.donghaienis, about 660 bp of ITS was amplif ied using primers of ITS1 (forward,5ʹ-TCCGTAGGTGAACCTGCGG-3ʹ) and ITS4(reverse, 5ʹ-GCATATCAATAAGCGGAGGA-3ʹ)(White et al., 1990). Polymerase chain reaction (PCR)reactions were conducted using a PCR Master Cycler nexus gradient (Eppendorf, Hamburg, Germany), and performed with a f inal volume of 25 μL, containing 9.5-μL ddH2O, 12.5-μL 2×Taq PCR MasterMix,1 μL of each PCR primer (10 mmol/L), and 1 μL of the DNA template. The following cycling conditions were used: an initial denaturation at 94 °C for 5 min, 35 cycles at 94 °C for 20 s, 55 °C for 30 s,and 72 °C for 2 min, and a f inal elongation step of 10 min at 72 °C. The PCR products were conf irmed using 1% agarose gel electrophoresis and visualized with ultraviolet light. Targeted bands were purif ied using an agarose gel DNA fragment recovery kit(GENEray Biotechnology, Shanghai, China), ligated with pMD18-T cloning vector (TaKaRa, Tokyo,Japan), and then sequenced (Sangon, Shanghai,China). Sequences were deposited in GenBank with accession numbers OL355144–OL355148 (LSU,B.brevisulcata), OL355142 (LSU,Bps.adriatica),OL314541 (LSU,S.donghaienis), and OL314542(ITS,S.donghaienis).
For surface sediment sample (S01-1), the single cysts were individually micropipetted onto a glass slide, then photographed with an inverted microscope(IX73, Olympus, Japan) equipped with a DP80 digital camera (Olympus, Japan). Subsequently, individual cyst was micropipetted and transferred onto a sterile slide, then broken by another coverslip. The crushed cyst and the coverslip pieces were transferred into a 250-μL centrifuge tube, as the template to amplify about 1 400 bp of the LSU rDNA using the primer set, D1R(forward, 5ʹ-ACCCGCTGAATTTAAGCATA-3ʹ)(Scholin et al., 1994) and 28-1483R (reverse,5ʹ-GCTACTACCACCAAGATCTGC-3ʹ) (Daugbjerg et al., 2000). The following procedure was according to Shang et al. (2019).
For phylogenetic analyses of the LSU rDNA regions ofB.brevisulcataandBps.adriatica, and LSU and ITS rDNA regions ofS.donghaienis,newly obtained LSU rDNA and ITS sequences were incorporated into those of closely related species available in the GenBank and that of outgroup taxa were f irst aligned using MAFFT v7.475 (Katoh et al.,2002) online program (http://maff t.cbrc.jp/alignment/server/) with default settings, and alignments were manually checked with BioEdit v7.2.5 (Hall, 1999).The f inal alignments of the LSU and ITS rDNA sequences ofS.donghaienisconsisted of 97 and 41 taxa and contained 750 and 600 positions (including gaps introduced from alignment), and the sequences ofCryptoperidiniopsisbrodyi(DQ991374) andPentapharsodiniumdalei(JX262496) were used as the outgroup, respectively. LSU rDNA sequences ofBps.adriaticaandB.brevisulcataconsisted of 66 taxa with 1 590 positions (including gaps introduced from alignment), and the sequence ofAlexandriummargalef ii(AY154957) was used as outgroup. The program jModelTest 2.1.4 was used to select the most appropriate model of molecular evolution with Akaike information criterion (AICc) (Posada, 2008),models GTR+G+I and TrN+I+G were selected as the best-f it model for the LSU rDNA and ITS datasets ofS.donghaienis, and TrN+I+G for the LSU rDNA dataset ofBps.adriaticaandB.brevisulcata.Phylogenetic trees were constructed using Bayesian inference (BI) and maximum likelihood (ML)analyses. Bayesian inference (BI) was performed with MrBayes 3.2.6 (Ronquist and Huelsenbeck,2003) with the best-f itting substitution models(GTR+I+G for LSU rDNA dataset ofS.donghaienis,TrN+I+G for ITS dataset ofS.donghaienis, and TrN+I+G for LSU rDNA dataset ofBps.adriaticaandB.brevisulcata). Four independent Markov chain Monte Carlo simulations were run simultaneously for 5 000 000 generations and trees were sampled every 1 000 generations. The f irst 10% trees were discarded as burn-in. The convergence was judged based on the average standard deviation of split frequencies(all less than 0.01). The remaining trees were used to generate a consensus tree and calculate the posterior probabilities of all branches using a majority-rule consensus approach. Maximum likelihood (ML)analyses were conducted with raxmlGUI v1.3.1(Silvestro and Michalak, 2012; Stamatakis, 2014)using the models GTR+I+G (for LSU rDNA dataset ofS.donghaienis), GTR+G (for ITS dataset ofS.donghaienis; the model GTR+G ranked the third, and the score of this model was close to model TrN+I+G),and GTR+I+G (for LSU rDNA dataset ofBps.adriaticaandB.brevisulcata; the model GTR+I+G ranked the second, and the score of this model was close to model TrN+I+G). Node support was assessed with 1 000 bootstrap replicates. FigTree (v1.4.4) was used to view and edit trees for publication.

Fig.1 Light microscopy photographs of Bps. adriatica and S. donghaienis cysts
2.7 Genetic diversity analyses
The pairwise distances were computed among all sequences that were newly obtained in the present work forB.brevisulcata,Bps.adriatica, andS.donghaienisand that retrieved from the NCBI database for these three species together with other reference sequences. Sequences were aligned using the MAFFT v7.475 with the default settings (Katoh et al., 2002) (http://maff t.cbrc.jp/alignment/server/) and modif ied manually using BioEdit v7.2.5 (Hall, 1999).Pairwise evolutionary distances were then computed using Jukes and Cantor algorithm implemented in the MEGA X (Tamura et al., 2004; Kumar et al., 2018).
3.1 Morphological observations of the resting cysts
Resting cyst ofBps.adriaticawas sub-spherical to spherical and light brown, with a diameter of ~7.3 μm,full of small granules and had 2–3 red accumulation bodies (Fig.1a). After four days’ incubation, the cyst was germinated. The cyst wall was thick and smooth,and the archeopyle was tremic (Fig.1b). Resting cyst ofS.donghaieniswas noncalcareous, spherical and brown, full of diff erent sizes of granules (Fig.1c). The diameter of the cyst was ~10.7 μm, and it contained several red accumulation bodies (Fig.1c). After four days’ incubation, the cyst was germinated. The cyst wall was thick, and the archeopyle was unclear(Fig.1d). As the f ive clonal cultures ofB.brevisulcatawere established from cyst assemblage germination,the resting and empty cysts of this species were not observed.
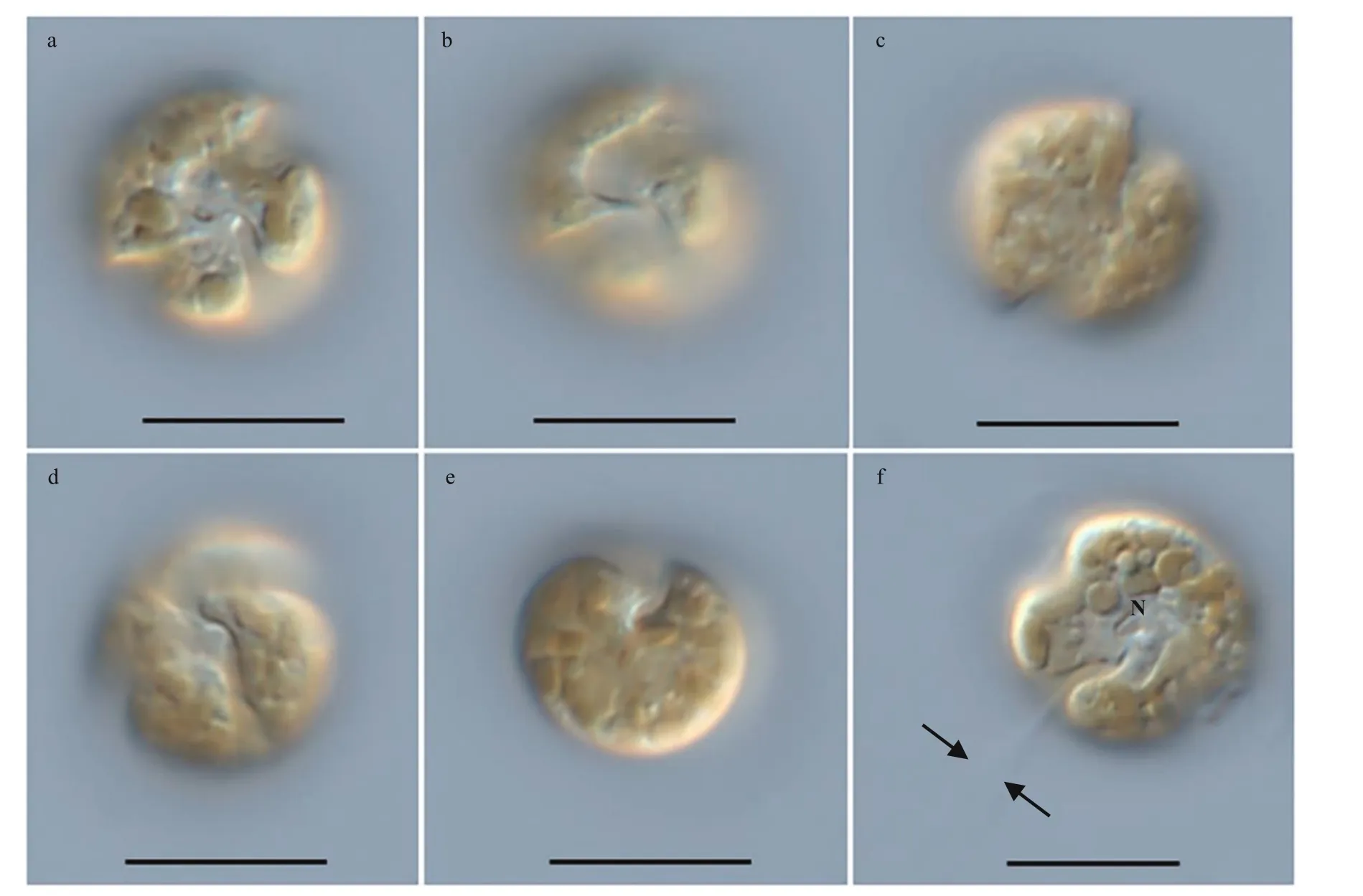
Fig.2 Light microscopy photographs of B. brevisulcata strain S1 germinated from resting cyst
3.2 Morphological observations for the vegetative cells of Biecheleria brevisulcata, Biecheleriopsis adriatica, and Scrippsiella donghaienis
The vegetative cells ofB.brevisulcatawere spherical to ellipsoidal, 7.2–9.2 μm long (average 8.2±0.6 μm;n=50) and 5.8–8.9 μm wide (average 7.3±0.7 μm;n=50) (Figs.2–3). The epicone was slightly wider and longer than the hypocone, and mushroom-shaped (Figs.2–3). The hypocone was bilobed (Figs.2–3). The descending cingulum was deep and wide, and displaced by 1.5 times its own width (Figs.2a–b, 3a–b). The sulcus was in the form of a sigmoid curve (Figs.2a, b, d, & 3a). The nucleus was round and located in the middle or slightly upper part of the hypocone (Fig.2f). Numerous yellowbrownish and reticulated or granulated chloroplasts were distributed peripherally (Fig.2). The arrangement of polygonal amphiesmal vesicles (AVs) was shown in Fig.3. Latitudinal rows of AVs were in four series on the epicone (E1–E4), three series on the hypocone(H1–H3), and three series in the cingular area (C1–C3; Fig.3b, d & e). A narrow elongate apical vesicle(EAV) was present in the apical area (Fig.3e–f), which was surrounded by f ive AVs (a small four-sided AV(X) and four elongated quadrangular AVs) and eight irregular pentagonal AVs (E1-1–E1-8; Fig.3e–f).
The vegetative cells ofBps.adriaticawas spherical to ellipsoid, 5.8–10.1 μm in length (average 7.8±1.2 μm;n=50) and 4.1–8.0 μm in width (average 6.0±0.9 μm;n=50). The epicone was rounded,and almost equal to the hypocone (Figs.4–5). The cingulum was deeper and wider than the sulcus(Figs.4–5). The cingulum was median and displaced by 1.5 times its own width (Figs.4–5). The sulcus was slightly sigmoid (Figs.4a–b, e–f, & 5a). Chloroplasts were yellowish-brown, and reticulated or granulated,which were distributed peripherally (Fig.4). The round refractive bodies were commonly observed(Fig.4). The nucleus was located in the center or slightly upper part of the hypocone (Fig.4c). Many pentagonal or hexagonal AVs were observed on the cell surface (Fig.5). Four epiconal, three cingular, and four hypoconal AV series were formed the latitudinal series (Fig.5a & c). The apical furrow was composed of an EAV and several surrounding AVs (Fig.5a–b, d,& f).
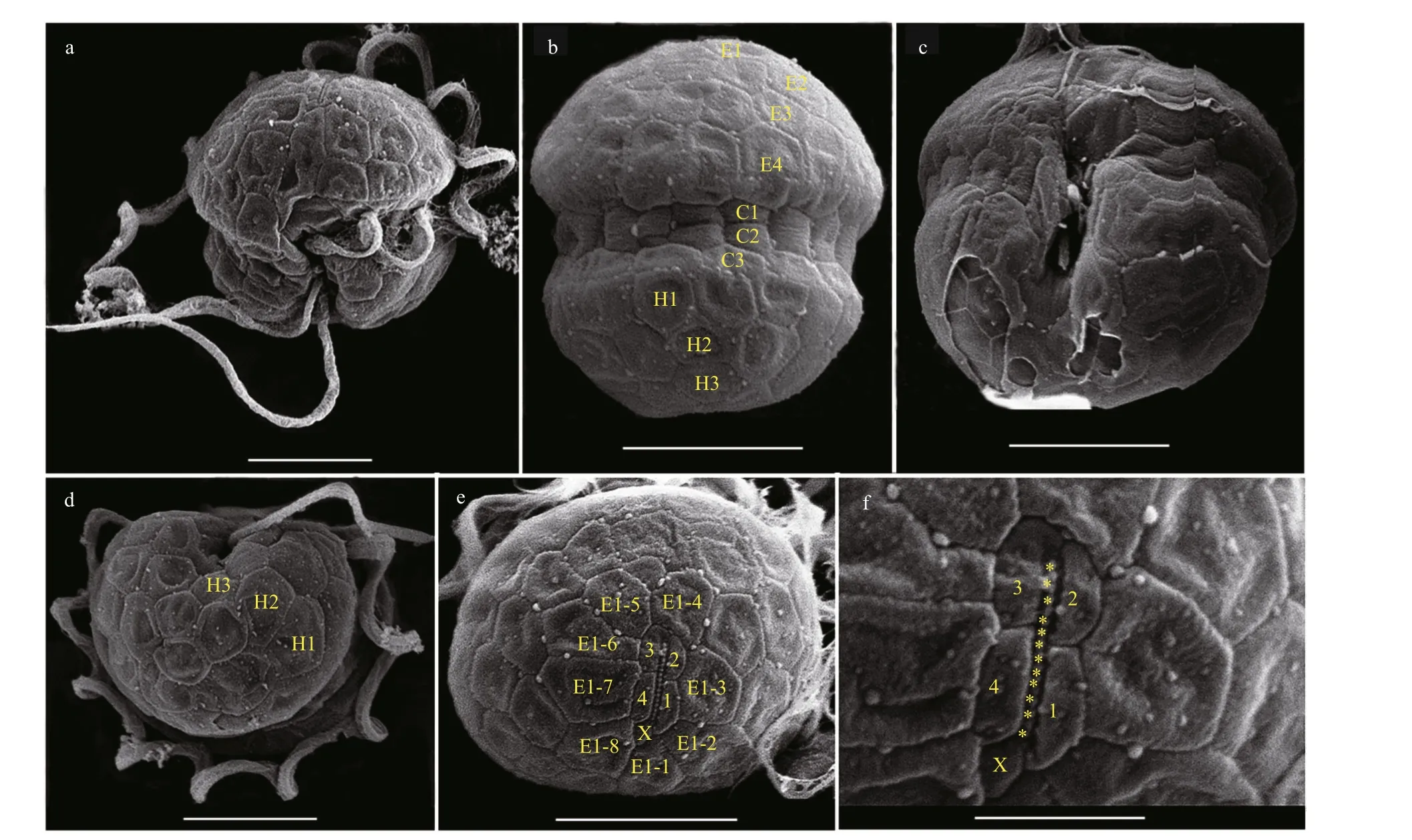
Fig.3 Scanning electron micrographs (SEM) of B. brevisulcata strain S1 germinated from resting cyst
The vegetative cells ofS.donghaieniswas 12.8–20.2 μm long (average 16.4±2.1 μm;n=50) and 9.6–16.0 μm wide (average 12.9±1.5 μm;n=50). The epitheca was conical and longer than the hypotheca,and the hypotheca was rounded and bilobed (Figs.6–7). The plate formula is Po, x, 4ʹ, 3a, 7″, 6c, 6s, 5‴, 2■(Fig.7). The cingulum was wide and deep (Fig.7a–c).The upper part of sulcus was narrower than its lower part (Figs.6a–b & 7a). The apical pore complex (APC)comprised a round apical pore plate and a long canal plate (Fig.7a & e–f). The nucleus was rounded and located centrally (Fig.6d). The yellowish-brown, and granulated chloroplasts were distributed peripherally(Fig.6d–e).
3.3 Pigment composition
Based on available standards, four photosynthetic pigments were identif ied inB. brevisulcata(strain S1),Bps.adriatica(strain S21), andS.donghaienis(strain 23), including one kind of chlorophyll (Chla)and three carotenoids (peridinin, diadinoxanthin, and diatoxanthin; Fig.8). Chl-acontents ofB.brevisulcata(strain S1),Bps.adriatica(strain S21), andS.donghaienis(S23) were 1.02, 0.25, and 10.89 pg/cell, respectively. Peridinin (3.51, 4.91, and 79.11 pg/cell) was the most abundant carotenoid for the three species, then diadinoxanthin, and diatoxanthin.There were 10, 11, and 11 unidentif ied small peaks(either new pigments or known pigments but without standards) forB.brevisulcatastrain S1,Bps.adriaticastrain S21, andS.donghaienisstrain S23.
3.4 Molecular phylogeny
Seven partial LSU rRNA gene sequences ofB.brevisulcata(strains S1, S2, S3, S4, and S5;OL355144–OL355148),Bps.adriatica(strain S21; OL355142), andS.donghaienis(strain 23;OL314541), and one ITS sequence ofS.donghaienis(strain 23; OL314542) were obtained from the clonal cultures. One partial LSU rRNA gene sequence ofBps.adriatica(ON350794) was obtained using single-cell PCR sequencing for the cyst from surface sediment sample of S01-1.
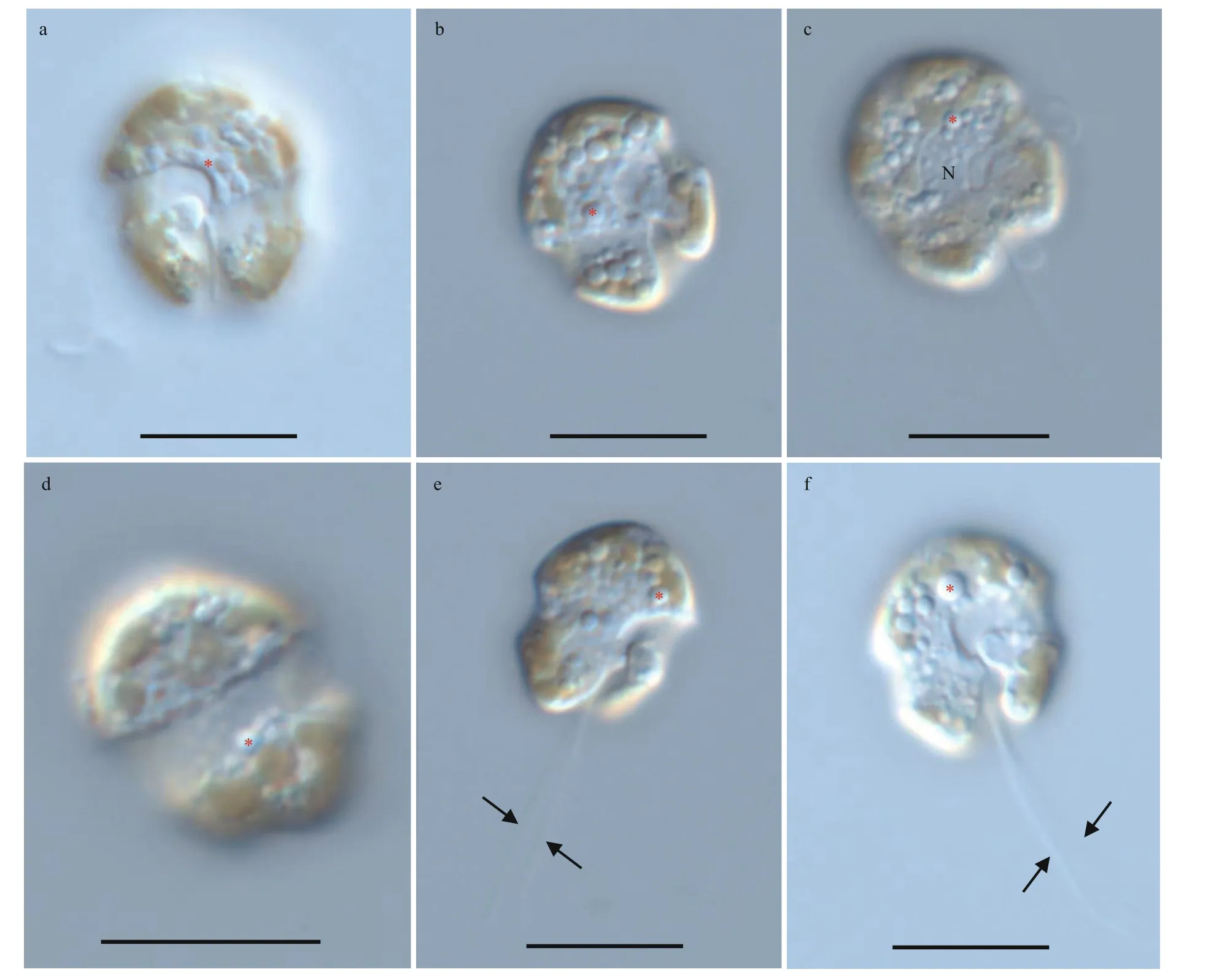
Fig.4 Light microscopy photographs of Bps. adriatica strain S21 germinated from resting cyst
The partial LSU rRNA gene sequence ofB.brevisulcatastrain S1 (1 374 bp; OL355144)was 99.93% (1 373 bp/1 374 bp) identical to the entity of the type material (AB858351), 99.88%(852 bp/853 bp) to 99.93% (1 359 bp/1 360 bp)identical to four entities (AB858352, AB858353,LC068842, and OL699922) deposited asB.brevisulcata, and 98.60% (845 bp/857 bp), 99.41%(509 bp/512 bp), 99.30% (853 bp/859 bp), and 99.32%(877 bp/883 bp) identical toB.pseudopalustris(syn.Woloszynskiapseudopalustris; AF260402),B.baltica(syn.Woloszynskia halophila sensuKremp et al.(2005); AY628430),B.cincta(syn.Woloszynskiacincta; FJ024705), andB.tirezensis(LT601379).
The partial LSU rRNA gene sequence ofBps.adriaticastrain S21 (1 425 bp; OL355142) was 99.69% (1 267 bp/1 271 bp) identical to the entity of the type material deposited at GenBank asGymnodiniumpygmaeumstrain K-0968, 99.24% (1 172 bp/1 181 bp)to 99.79% (1 422 bp/1 425 bp) identical to 12 entities(AB858354–AB858356, LC068843, LC413947–LC413950, LM992904–LM992906, and OL691545)deposited asBps.adriatica, 99.78% identical to 11 entities (KM603188–KM603198) deposited asBps. cf.adriatica, and 99.13% (794 bp/801 bp) to 99.30% (1 415 bp/1 425 bp) identical to four entities(JN558103–JN558105, KM603185) deposited asProtodiniumsimplex. Among all these entities in GenBank,Bps.adriaticaalso corresponded to the cultures established from vegetative cells isolated from the same area where the cultures from cysts were established (i.e., East China Sea; Luo et al., 2015).
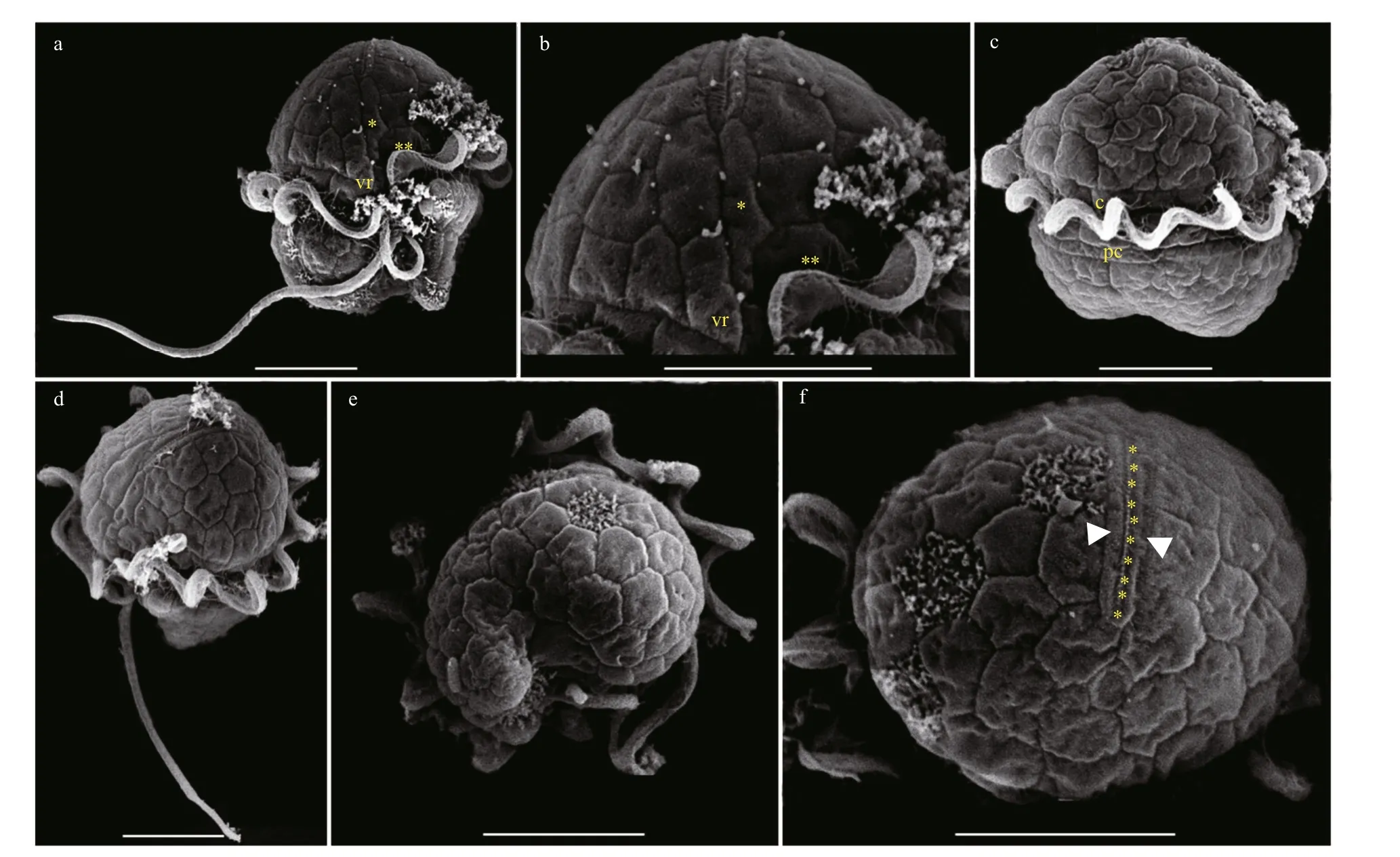
Fig.5 Scanning electron micrographs (SEM) of Bps. adriatica strain S21 germinated from resting cyst
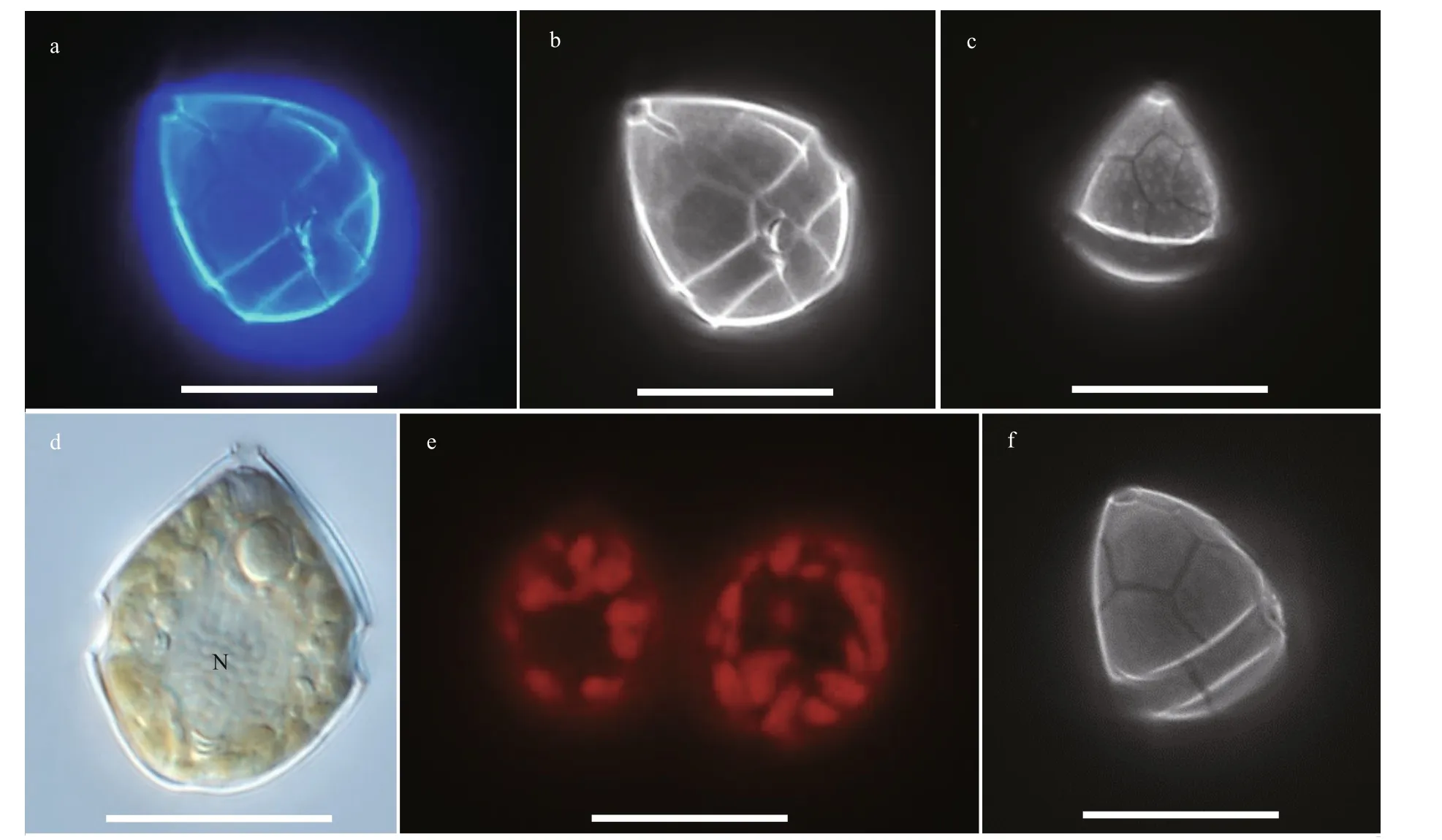
Fig.6 Light microscopy photographs of S. donghaienis strain S23 germinated from resting cyst
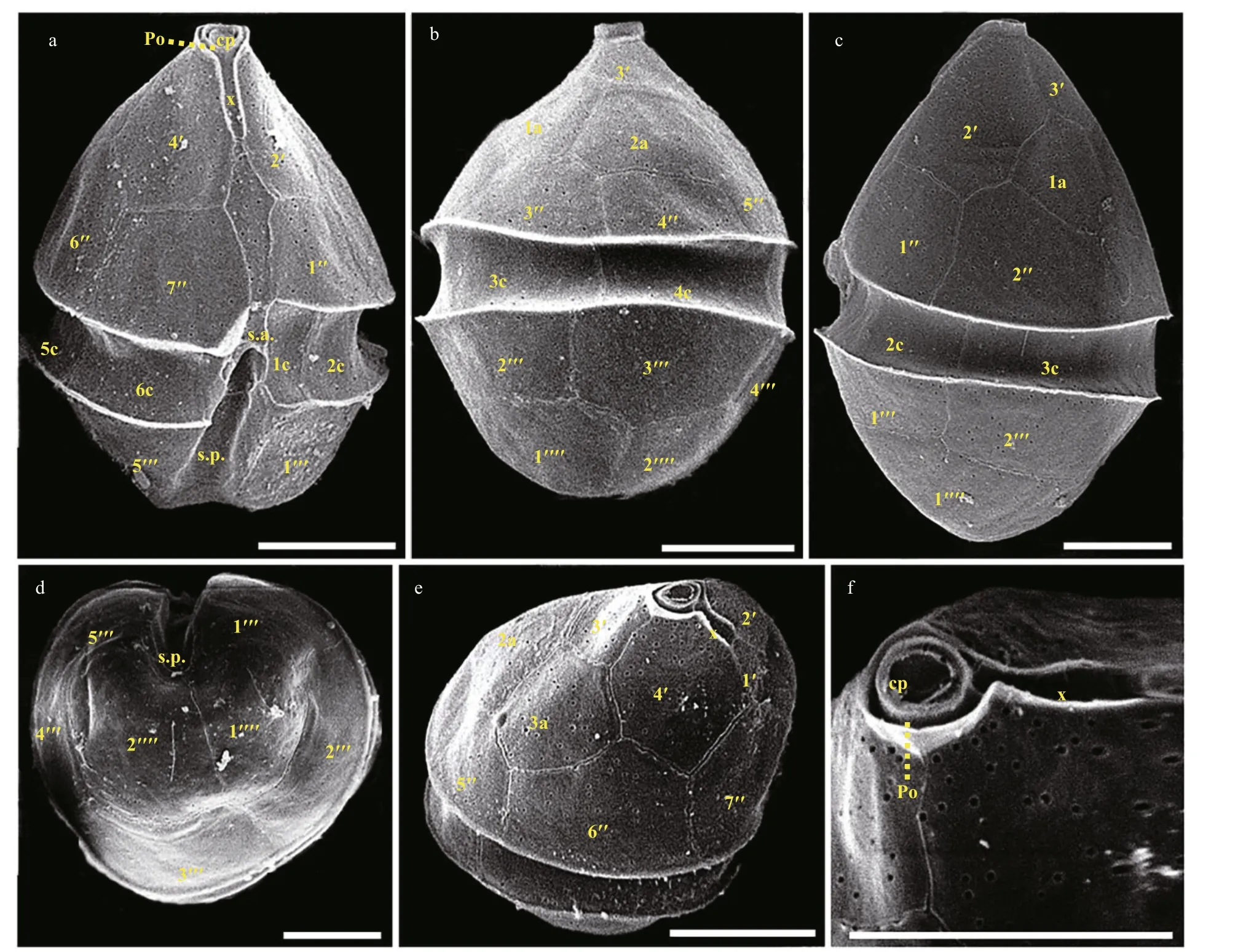
Fig.7 Scanning electron micrographs (SEM) of S. donghaienis strain S23 germinated from resting cyst
The partial LSU rRNA gene sequence ofS.donghaienisstrain 23 (1 433 bp; OL314541) was 98.51% (796 bp/808 bp) to 99.51% (812 bp/816 bp)identical to 69 entities deposited asS.donghaienisin GenBank. The ITS sequence ofS.donghaienisstrain 23 (663 bp; OL314542) was 93.80% (454 bp/484 bp)to 99.64% (558 bp/560 bp) identical to f ive entities(AY685008, HQ729492, HQ729502, JN982374, and MG914024) deposited asS.donghaienis, and 99.28%(550 bp/554 bp) to 99.68% (617 bp/619 bp) identical to seven entities (AY499533, AY676151, AY67615,AY788357, and FJ823594–FJ823596) deposited asScrippsiellasp. in GenBank.
Phylogenetic analyses ofB.brevisulcata,Bps.adriatica, andS.donghaienisusing maximum likelihood (ML) and Bayesian inference (BI)generated similar trees based on LSU rRNA gene and ITS sequences but diff ered at a few internal nodes(Figs.9–11). ForB.brevisulcata, our sequences and other sequences (AB858351–AB858351,LC068842) formed a coherent clade with strong support (0.71/100), which is sistering to the clade includingWoloszynskiahalophila(EF205019,AY628430),B.cincta(FJ024705),Woloszynskiapseudopalustris(AF260402), andGymnodiniumsp. (AY318248) with strong support (0.95/100;Fig.9). ForBps.adriatica, our sequence and other sequences (LC068843, LC413947, and LC413948),Bps. cf.adriatica(KM603188–KM603198), andG.corii(GU477610) formed a coherent clade with strong support (0.93/100; Fig.9), and formed a wellsupported sister clade (0.98/100; Fig.9) including the type material ofBps.adriatica(“G.pygmaeum”,EU857537), andG.corii(EU165298, AF318226).For the phylogenetic analysis ofS.donghaienisbased on LSU rRNA gene sequences, our sequence(OL31454), 69 sequences deposited asS.donghaienis,andScrippsiellasp. (AY685011) formed a coherent clade with maximal support (1/100; Fig.10),and formed sister groups with otherScrippsiellaspecies (Fig.10). For the phylogenetic analysis ofS.donghaienisbased on ITS sequences, our sequence (OL314542), four sequences deposited asS.donghaienis(HQ729492, HQ729502, JN982374,and MG914024), and seven sequences deposited asScrippsiellasp. (AY499533, AY676151, AY676155,AY788357, FJ823594–FJ823596) formed a coherent clade with medium support (0.51/88; Fig.11), and formed a sister group with one sequence deposited asS.donghaienis(AY685008) with maximal support(1/100; Fig.11).
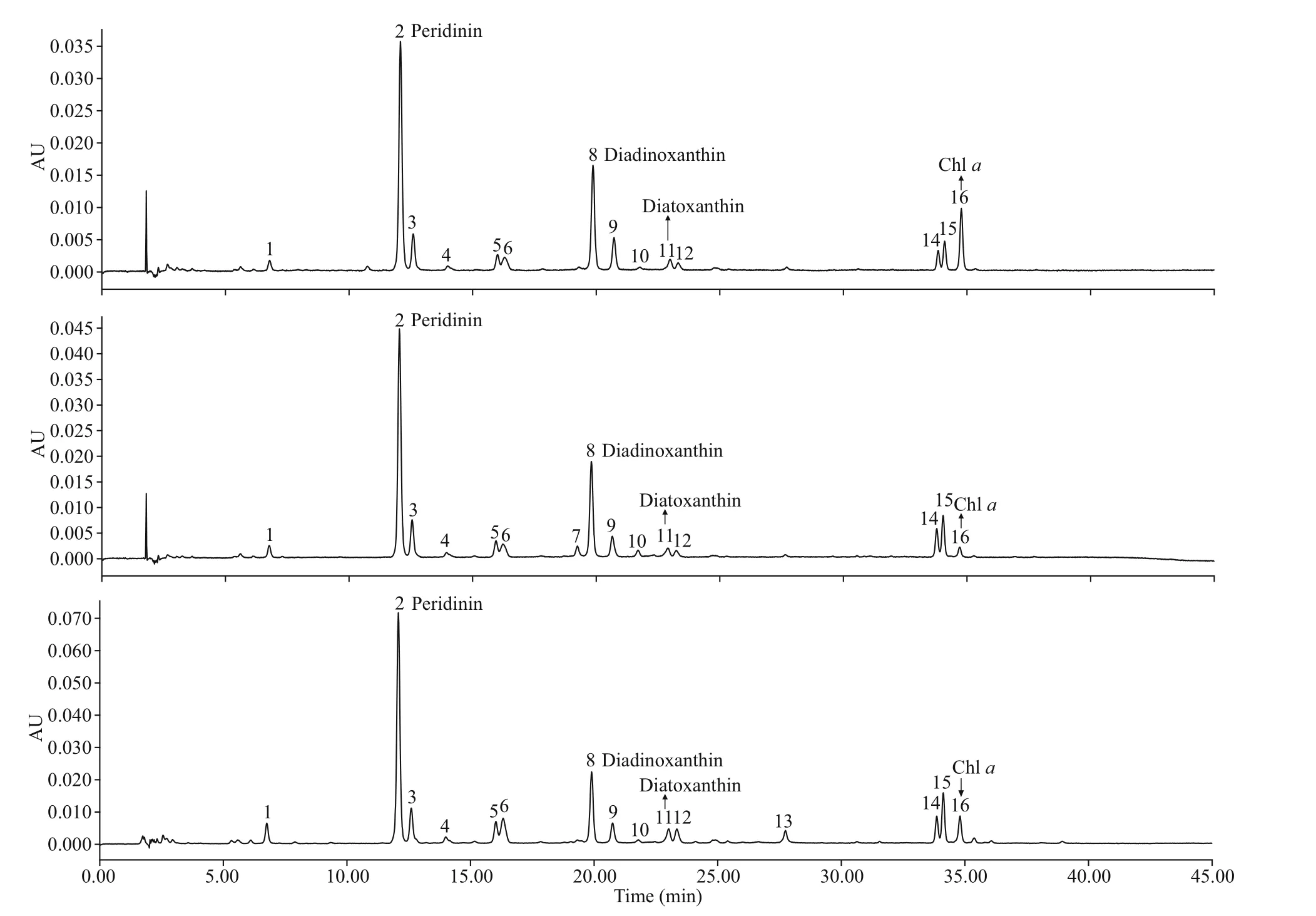
Fig.8 Absorption chromatograms (440 nm) of the pigment extracts of B . brevisulcata strain S1, Bps. adriatica strain S21, and S. donghaienis strain 23 from the East China Sea, China
3.5 The genetic diversity ref lected in partial rDNA and ITS sequences
The pairwise distances computed using Jukes-Cantor model showed that the sequence divergence among the LSU rRNA gene sequences ofBps.adriaticafrom the present study, type material(deposited in GenBank asG.pygmaeumwith the accession No. EU857537), and sequences deposited asBps.adriatica(AB858354–AB858356, LC068843,LC413947–LC413950, and LM992904–LM992906)ranged from 0.000 to 0.006, the distances between our sequence andBps. cf.adriatica(KM603188–KM603198) were 0.002, and among our sequence, and the sequences deposited asP.simplex(AF060901),Gymnodiniumsp. (EF205006),G.corii(AF318226,EU165298, GU477610, KT389967, KT389942),P.simplex(FJ024704, JN558103–JN558105,KM603185–KM603187) ranged from 0.000 to 0.007 (Supplementary Table S1). However, the distances between our sequence andB.brevisulcata(AB858351),Ansanellagranifera(HG529980),andA.margalef ii(AY154957) were 0.039, 0.029,and 0.244, respectively (Supplementary Table S1).Sequence diff erences among our strain (S21), the other stains from water samples or surface sediments formed a well-supported clade with ours (Fig.9), and the one we found in the surface sediment sample in the same sea area were found mainly at three stable positions (3 bp/1 438 bp (D1–D6 regions); Fig.12).
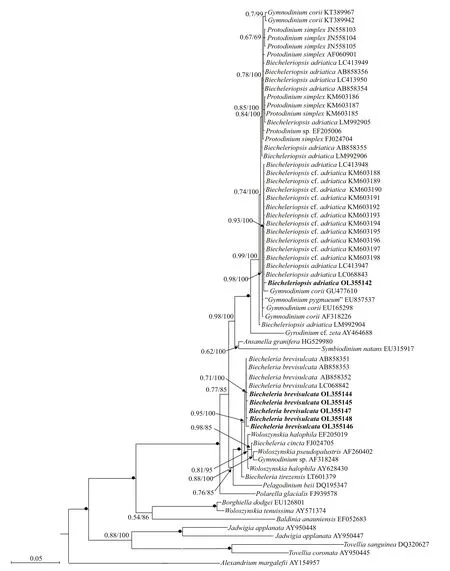
Fig.9 Molecular phylogeny of B. brevisulcata, Bps. adriatica, and diverse assemblage of other dinof lagellates inferred from the partial LSU rRNA gene sequences based on Bayesian inference (BI) with A. margalef ii (AY154957) as outgroup

Fig.10 Molecular phylogeny of S. donghaienis and diverse assemblage of other dinof lagellates inferred from the partial LSU rRNA gene sequences based on Bayesian inference (BI) with C. brodyi (DQ991374) as outgroup
The genetic distance based on LSU rRNA gene ofB.brevisulcataobtained in the present study and previous works, and other related species were compared (Supplementary Table S2). The sequence divergence among the LSU rRNA gene sequences ofB.brevisulcataobtained in our work (OL355144–OL355148) and other strains previously deposited in GenBank (AB858351–AB858353, LC068842)ranged within 0.000–0.007 (Supplementary Table S2), the sequence divergence amongB.brevisulcata(OL355144–OL355148) and otherBiecheleriaspecies (B.baltica,B.cincta,B.pseudopalustris, andB.tirezensis) ranged 0.002–0.015 (Supplementary Table S2), but the sequence divergence amongB.brevisulcata(OL355144–OL355148) and other distant speciesPelagodiniumbeii(DQ195347),Polarellaglacialis(FJ939578), andA.margalef ii(AY154957) were 0.055–0.057, 0.077–0.079, and 0.808–0.875, respectively (Supplementary Table S2).
The genetic distance based on LSU rRNA gene and ITS ofS.donghaienisobtained in the present study and previous work, and other related species were compared(Supplementary Tables S3 & S4). The sequence divergence among the LSU rRNA gene sequence ofS.donghaienisobtained in our work (OL314541) and other strains previously deposited in GenBank ranged 0.000–0.009 (Supplementary Table S3), 0.013–0.139,and the sequence divergence amongS.donghaienis(OL314541) and otherScrippsiellaspecies (S.acuminate,S.erinaceus,S.sweeneyae,S.spinifera,S.plana,S.bicarinata,S.kirschiae,S.trif ida,S.infula,S.rotunda,S.lachrymose,S.enormis,S.masanensis,S.precaria, andS.ramonii) ranged 0.002–0.015(Supplementary Table S3). The pairwise distances computed using Jukes-Cantor model showed that the sequence divergence between the ITS sequences ofS.donghaienisobtained in the present study, and sequences deposited asS.donghaienis(AY685008,HQ729492, HQ729502, JN982374, and MG914024)ranged from 0.000 to 0.015 (Supplementary Table S4),the sequence divergence between the ITS sequences ofS.donghaienisobtained in the present study, and the otherScrippsiellaspecies ranged 0.175–0.360(Supplementary Table S4). Sequence diff erences between our strain (S23) and the other stains from water samples or surface sediments formed a wellsupported clade with ours (Fig.11) were found mainly at two stable positions (2 bp/599 bp; Fig.13).
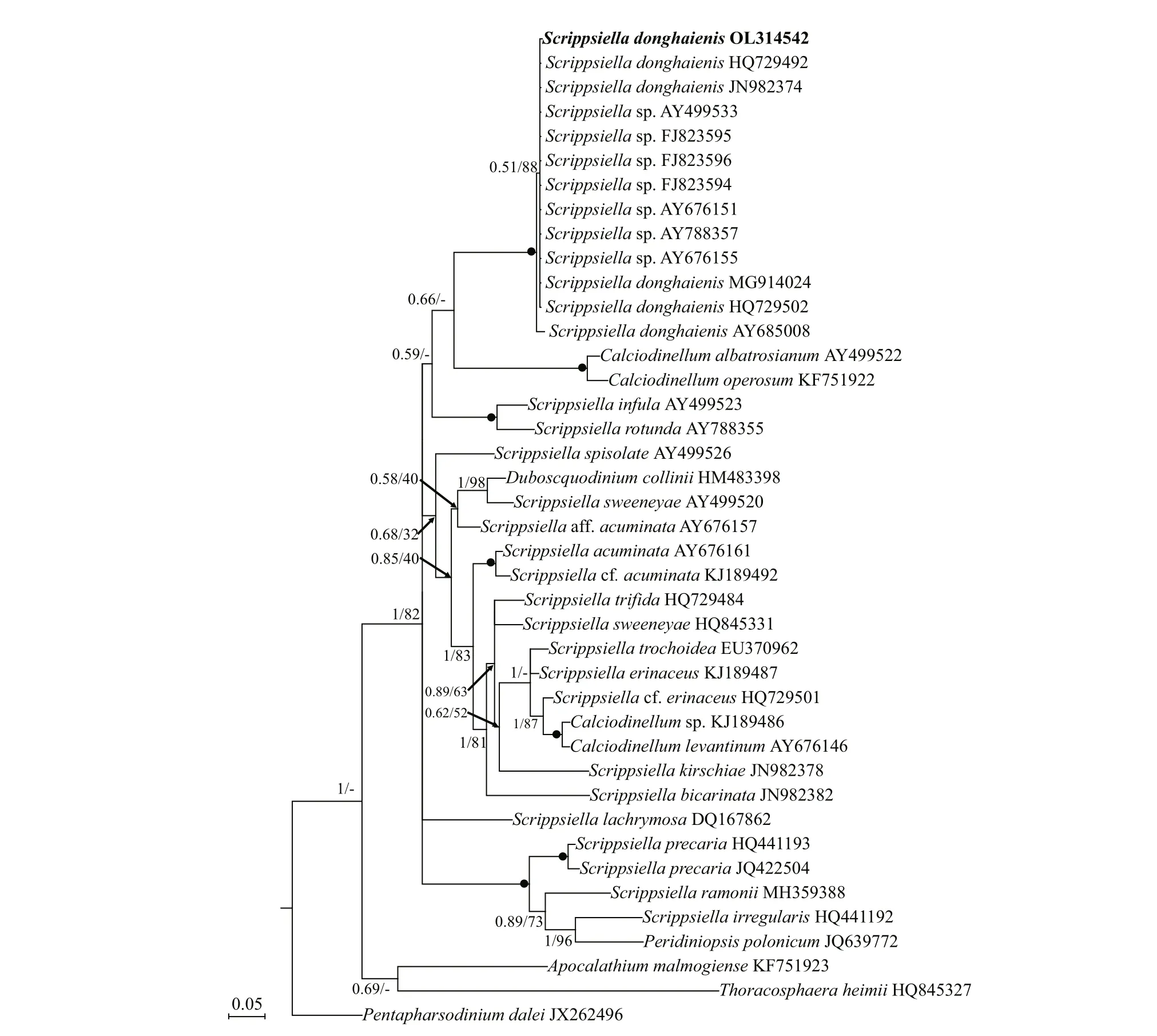
Fig.11 Molecular phylogeny of S. donghaienis and diverse assemblage of other dinof lagellates inferred from the internal transcribed spacer (ITS) region sequences based on Bayesian inference (BI) with P. dalei (DQ991374) as outgroup
4.1 Identif ication of Biecheleriopsis adriatica resting cyst presented in the f ield sediment

Fig.12 Sequence variation in the LSU rDNA sequences among Bps. adriatica strain 21, the one cyst of this species found in the surface sediment sample in the same sea area, and other strains of Bps. adriatica formed a well-supported clade with ours in Fig.9
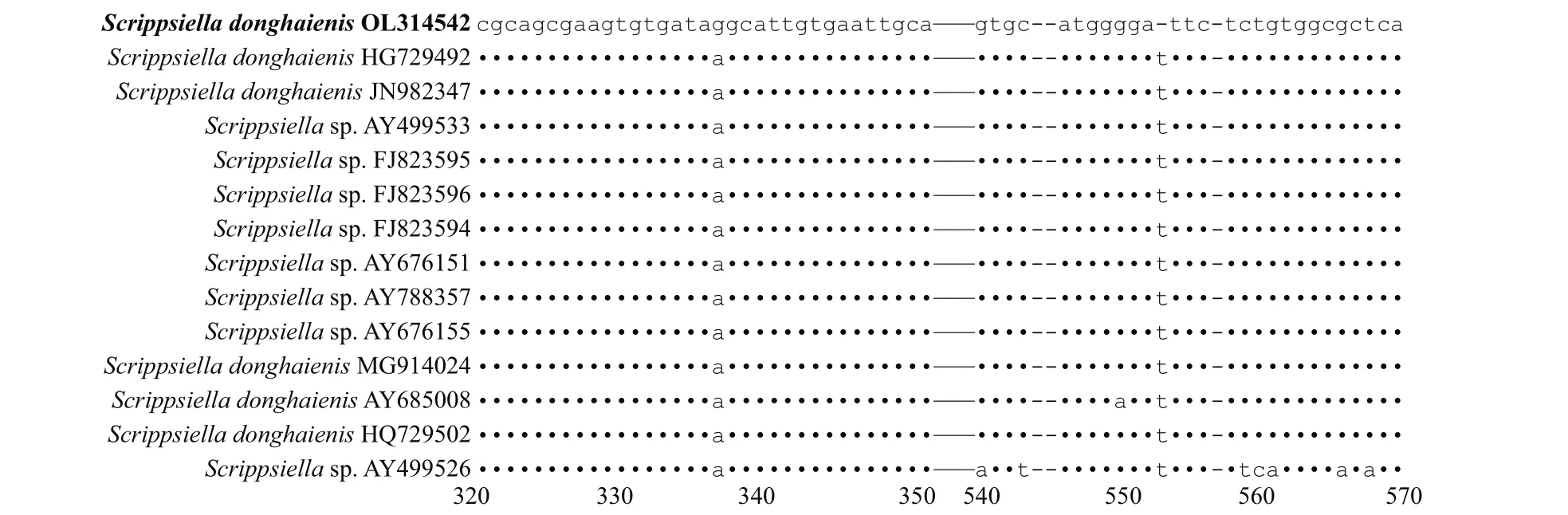
Fig.13 Sequence variation in the ITS sequences between S. donghaienis strain 23 and the other strains formed a well clade with ours in Fig.11
Biecheleriopsisadriaticawas described by Moestrup et al. (2009) who diff erentiated it from the species ofBiecheleriabased on the presence of a nuclear connector and a 51-bases long fragment of D2 domain of LSU rRNA gene. Moestrup et al. (2009)also found resting cysts in the culture ofBps.adriatica,described as having ellipsoidal to ovoid shapes and numerous spines, a size of 7–8-μm length and 5–6-μm width. Benico et al. (2019) found resting cyst-like cells in unialgal culture being morphologically diff erent from vegetative cell, and they called them as resting cyst-like cells, which were ~16 μm, spherical,covered by a transparent thick wall, and contained a red accumulation body. Kang et al. (2009), Wang et al. (2013), and Kang and Wang (2018) germinatedBps.adriaticafrom cyst assemblages (identif ied asG.coriiin Kang et al. (2009) and Wang et al. (2013))from Southern Chinese coastal sediments, but did not observe the genuine resting cyst of this organism.Luo et al. (2015) also germinatedBps. cf.adriatica(very possible conspecif ic with our strain S21) from the sediment collected from the Yellow Sea and the South China Sea, but also without observation on the morphology ofBps. cf.adriaticacyst. The abovementioned works onBps.adriaticahave conf irmed that it could form resting cyst, but none of them reported the morphology of cyst from the f ield. In this work, we germinatedBps.adriaticafrom a single cyst collected from the East China Sea, and the species was identif ied with morphological and molecular characterization allowing us to conf idently conf irm the cyst-motile stage relationship of this small-sized species. Resting cyst ofBps.adriaticawas spherical and sub-spherical, which is similar to the so-called“resting cyst-like cells” as observed by Benico et al.(2019), but diff erent from the cyst from culture having an ellipsoidal to ovoid morphology (Moestrup et al.,2009). The diameter ofBps.adriaticain our work(~7.3 μm) is very close to the cyst (7–8 μm long and 5–6 μm wide) observed by Moestrup et al. (2009), but much smaller than the resting cyst-like cells (~16 μm)observed by Benico et al. (2019). The sieves with the pore size larger than 20 μm were routinely used to concentrate cysts, and then followed by cyst assemblage germination, the cysts with smaller size(<20 μm) would be lost during processing, therefore,the eff orts in discovering more small sized cysts should focus on the cyst assemblage being smaller than 20 μm.
4.2 Viability of dinof lagellates cysts stored in longburied sediments
In the past, changes of species composition in sediments have been used to assess environmental changes including eutrophication, changes in salinity, or oxygen concentration (Dale et al., 1999;Ellegaard et al., 2013, 2020; Bringué et al., 2016;García-Moreiras et al., 2018; Li et al., 2021; Siano et al., 2021). The adaptive responses of dinof lagellate species might be inf luenced by anthropogenic activities or climate changes, and revealing the evolutionary processes in a species is very important to understanding the genetic structure of populations.However, the length of live resting cyst preserved in the sediment cores (e.g., decades to centuries) or the molecular information (rRNA genes or other gene markers) for the buried cysts or residual fragments have not been well explored before 2010. After that, the impacts of environmental changes on the physiology, genetic structure, and diversity in various dinof lagellate species (Ribeiro et al., 2011; Klouch et al., 2016; Lundholm et al., 2017; Kremp et al.,2018; Delebecq et al., 2020; Ellegaard et al., 2020;Girault et al., 2021) have been investigated. In this study, we successfully revivedB.brevisulcata,Bps.adriatica, andS.donghaienisfrom a sediment core dated back to 1941±18 AD from the East China Sea,which indicates that these cysts are viable for at least 70 years. Klouch et al. (2016) detected molecular signal ofS.donghaienisin a sediment core sample dated back to 1866±7 AD from the Bay of Brest,France, and successfully germinated this organism only from the layers of 2–17 cm corresponding to the years of 2010±1 AD to 1978±2 AD, much more recent than 1941±18 AD. In older sediments (70–100 years),Protoceratiumreticulatum,Lingulodiniumpolyedrum, andP.daleiwere successfully germinated(Lundholm et al., 2011; Ribeiro et al., 2011). Girault et al. (2021) successfully germinatedA.minutumandS.acuminatafrom the sediment corresponding to 1947±11 AD. Kremp et al. (2018) foundApocalathiummalmogienseis viable in the sediment layers of 106-year old. Recently, Delebecq et al. (2020) germinatedA.minutum,Heterocapsaminima,Margalef idiniumpolykrikoides,Protoperidiniumspp., andDiplopsalisgroup in sediments dated back to 117±21 years ago,andS.acuminatabeyond 156±27 years ago. Our work increases the diversity of species (B.brevisulcataandBps.adriatica) potentially revivable from more longburied sediments, which will promote studies in the f ield of resurrection ecology.
4.3 Genetic diversity among dinof lagellates revived from cysts stored in long-buried sediments and current condition
Genetic diversity is an important aspect of biodiversity, which is def ined as measurements that determine the changes of genetic variability within any level of a taxon but more often within a species or even population, e.g., allelic diversity or richness,mutational diversity, and eff ective population size(Hughes et al., 2008; Ebenezer et al., 2012; Ellegren and Galtier, 2016). Previous phylogeographic studies have revealed high levels of genetic diversity in a number of dinof lagellate species, e.g.,A.fundyense(Erdner et al., 2011),Amphidiniumspp. (Murray et al., 2012),Gambierdiscusspp. (Nishimura et al., 2013),Margalef idiniumfulvescens(Lin et al.,2020),Ostreopsisspp. (Lee and Park, 2020), andPseudocochlodiniumprofundisulcus(Hu et al., 2021).However, all above-mentioned works examined the genetic diversity among contemporary populations or even within a population rather than that between historical and contemporary populations as done in the present work, although the history here is less than a century (~70 years). From our phylogenetic tree, our f ive strains ofB.brevisulcatabranched together with the other strains of this species, but their evolution distance varied. Our strain ofBps.adriaticaand three strains ofBps.adriatica, eleven strains ofBps. cf.adriatica, and one strain ofG.coriiformed a coherent clade, and a sister group with the type material of this species, but diff ered at three stable positions. Our strain ofS.donghaienis, the other strains of this species, and one strain ofScrippsiellasp. formed a coherent clade. Within this clade, there were four branches, our strain grouped together with those historical and contemporary populations.We also compared the rRNA gene based genetic distances of the seven strains ofB.brevisulcata,Bps.adriatica, andS.donghaienisrevived from a long-buried sediment dated back to 1941±18 AD and those from water samples and surface sediments (the contemporary age), and found the genetic distance between our strain ofB.brevisulcataand other strains range from 0.002 to 0.006, but among other strains,the genetic distances are 0.000–0.005, most 0.001,and similar trends were also found inBps.adriatica,andS.donghaienis. We also found diff erences in three stable positions of LSU rRNA gene sequences between the population ofBps.adriaticawe found 70 years ago and the contemporary or present-day population (including the cyst of this species we found in the surface sediment and vegetative cells found by Luo et al. (2015) in the same sea area),and ITS sequences in two stable positions between the population ofS.donghaieniswe found 70 years ago and the contemporary or present-day population(including the cyst of this species found by Gu et al.(2008) in the surface sediment in the same sea area).It seemed these current populations are genetically diff erent from those that existed in the area 70 years ago, which suggests that there has been a shift in the populations ofBps.adriaticaandS.donghaienis.Due to a limited size of dataset for the present work,the possible historical succession of populations ofBps.adriatica,S.donghaienisand even other species in the area requires a more intensive and extensive investigation, as these possible shifts may be highly indicative of the environmental changes and anthropological activities that occurred in the area during the past.
All data generated and/or analyzed during the current study are available from the corresponding author on reasonable request.
We are highly grateful of the two anonymous reviewers for their patience, critical comments, and generous suggestions, which helped greatly the improvement of the manuscript. Sediment samples’collections were supported by NSFC Open Research Cruise, funded by Shiptime Sharing Project of NSFC.We appreciate for the cruises conducted by R/VXiangYangHong18and staff from First Institute of Oceanography, Ministry of Natural Resources, China.
- 范文大全
- 说说大全
- 学习资料
- 语录
- 生肖
- 解梦
- 十二星座
-
主题党日活动交流发言8篇
主题党日活动交流发言8篇主题党日活动交流发言篇13月13日,东城区党史学习教育动员大会召开。市委
【活动总结】 日期:2022-12-23
-
2022年4月主题党日活动记录范文15篇
2022年4月主题党日活动记录范文15篇2022年4月主题党日活动记录范文篇1一个崇尚阅读的民族,必然精神饱满、意气风发、活力四射。习近平总书记强调:“学习
【活动总结】 日期:2022-08-01
-
家乡赋|最美的家乡赋
家乡赋 孙传志 今安康市,白河双丰镇,吾之家乡也。三环沃土,山水环抱。其北依山,山系五岭,山
【调研报告】 日期:2020-04-01
-
少先队的光荣历史故事 队前教育-光辉历程
2017-2018学年队前教育1光辉历程一、劳动童子团1924——1927二、三十年代年的中国是一个
【法律文书】 日期:2020-06-23
-
党支部1-12月全年主题党日活动计划表
2022年党支部主题党日活动计划表序号活动时间活动方式活动内容12022年1月专题学习研讨集中观看2022年新年贺词,积极开展学习研讨交流。组织生活会组织党员认真对照党章...
【活动总结】 日期:2022-10-14
-
【人教版1-6年级数学上册知识点精编】1-6年级数学人教版教材
人教版二年级数学上册知识点汇总第一单元长度单位一、米和厘米1、测量物体的长度时,要用统一的标准去测量
【调研报告】 日期:2020-11-08
-
2022年2月份主题党日活动记录5篇
2022年2月份主题党日活动记录5篇2022年2月份主题党日活动记录篇1尊敬的党组织:在今年的开学初,本人积极参加教研室组织的教研活动,在学校教研员的指
【活动总结】 日期:2022-08-12
-
医院最佳主题党日活动11篇
医院最佳主题党日活动11篇医院最佳主题党日活动篇1 医院最佳主题党日活动篇2为隆重纪念中国共产党成立100周年,进一步巩固党的群众路线教育实践活动成果,切实
【活动总结】 日期:2022-10-29
-
2023年平安校园建设方案13篇
平安校园建设方案“平安校园”创建工作,我们幼儿园全体教职员工一直把它当作头等大事来抓。领导高度重视,以“平安校园”创建活动为抓手,建立和规范校园安全工作机制
【规章制度】 日期:2023-11-02
-
主题党日活动记录202210篇
主题党日活动记录202210篇主题党日活动记录2022篇12021年是中国共产党成立100周年,为广泛开展爱国主义宣传教育,铭记党的历史,讴歌党的光辉历程,
【活动总结】 日期:2022-08-02
-
正式的晚宴邀请函 公司晚宴邀请函
尊敬的先生 女士: 我公司谨定于xxxx年xx月xx日xx:xx在xxxx店隆重举行xx市xx届xxxx晚宴(宴会地址:xx区xx路xxxx) 敬请届时光临!xxxxxx集团股份有限公司xxxx有限公司敬邀xxxx年xx月xx日
【简历资料】 日期:2019-08-03
-
一年级新学期目标简短_一年级学生新学期打算
新学期到了,我是一年级下册的小学生了。 上课的时候,我要认真学习,不做小动作,认真听讲。我要认真学习,天天向上,努力学习,耳朵要听老师讲课,眼睛要瞪得大大的看老...
【简历资料】 日期:2019-10-26
-
《国行公祭,为佑世界和平》课文原文阅读_国行公祭为佑世界和平每段段意
国行公祭,为佑世界和平钟声“国行公祭,法立典章。铸兹宝鼎,祀我国殇。”侵华日军南京大屠杀遇难同胞纪念
【简历资料】 日期:2020-11-28
-
[信访复查复核制度作用探讨]信访复查复核有用吗
作为我国特有的一项制度,信访制度的出现并长期存在不是偶然的,虽然一些法学专家认为信访制度具有“人治”
【职场指南】 日期:2020-02-16
-
[党员干部2019年主题教育个人问题检视清单及整改措施2篇] 党员干部
2019年主题教育问题检视清单及整改措施根据主题教育领导小组办公室《关于认真做好主题教育检视问题整改
【求职简历】 日期:2019-11-08
-
网络维护工作内容_(精华)国家开放大学电大专科《网络系统管理与维护》形考任务1答案
国家开放大学电大专科《网络系统管理与维护》形考任务1答案形考任务1理解上网行为管理软件的功能【实训目
【职场指南】 日期:2020-07-17
-
红旗颂朗诵稿原文【《红旗颂》朗诵词】
《红旗颂》朗诵词 女:晴空万里,红旗飘扬, 六十载风云,我们昂首阔步。 男:六十个春秋,
【职场指南】 日期:2020-02-16
-
党委会与局长办公会的区别_局长办公会制度
为进一步加强xxx局工作的规范化、制度化建设,提高行政效能,规范议事程序,特制定本制度。一、会议形式1、局长办公会议由局长、副局长参加。由局长召集和主持。根据工作需要...
【求职简历】 日期:2019-07-30
-
《铁拳砸碎“黑警伞”》警示教育片观后感
影片深刻剖析了广西北海市公安局海西派出所原所长张枭杰蜕变堕落的轨迹。观看警示教育片后,做为一名党员教
【简历资料】 日期:2020-08-17
-
如何凝心聚力谋发展【坚定信心谋发展凝心聚力促跨越】
当前,清河正处于在苏北实现赶超跨越基础上全面腾飞的战略机遇期,处于在全市率先实现全面小康基础上率先实
【简历资料】 日期:2020-03-17
-
党建促乡村振兴现场会发言材料
现场指事件或行动发生的地点,也指接从事生产、工作、试验的场所,是事件或行动发生的地点,也是在需要行动的地点,在编程中,当前正在执行的程序指令,这是编程中的现场,在...
【其他范文】 日期:2022-09-12
-
企业:xx公司2020年双拥工件总结及2021年工作计划|社区双拥工作计划
XX公司2020年双拥工作总结及2021年工作计划2020年,XX公司及各成员企业双拥工作在集团公司
【毕业论文】 日期:2021-01-21
-
质检员工作总结汇报总结8篇:
质检员工作总结汇报总结8篇质检员工作总结汇报范文模板【一】20XX年,在我厂各领导的有力组织下,通过
【其他范文】 日期:2021-09-27
-
基层党建工作调研报告范文 [公司党建工作(非在职党员)调研报告]
公司党建工作(非在职党员)调研报告 为了积极探索加强我公司非在职党员管理工作的新途径,进一步摸清底
【节日庆典】 日期:2020-06-03
-
2022共青团入团时间_共青团入团申请模板【完整版】
当前位置:>>>2021-10-04敬爱的团支部:我是×年×班的×××,我通过班上团组织和同学的教育和帮助,我深深地认识到作为一个新世纪的青年,必须积极争取加入青年的组织----中...
【其他范文】 日期:2022-11-03
-
2022年度在省推动全民健身与全民健康深度融合会议上讲话
当前位置:>>>2022-03-17讲话,是指说话;发言;谈话,或者指演说和普及性的著作体裁。以下是好范文网分享的2022年在省推动全民健身与全民健康深度融合会议上的讲话,希望能帮...
【其他范文】 日期:2022-12-05
-
血清4型禽腺病毒Fiber-2蛋白截短表达及单克隆抗体制备
韦悠,谢芝勋,邓显文,李小凤,谢志勤,范晴,张艳芳,黄娇玲,王盛(广西兽医研究所 广西兽医生物技术重
【其他范文】 日期:2023-01-20
-
延续性护理模式对老年痴呆患者神经精神症状的影响
田雪萍弓敏(1 山阳县人民医院内科,陕西山阳726400;2 宝鸡市中心医院神经内科,陕西宝鸡721
【其他范文】 日期:2023-07-03
-
开展党员干部和公职人员涉酒违纪违法问题专项整治工作方案(全文)
开展党员干部和公职人员涉酒违纪违法问题专项整治工作方案根据中共xx市纪律检查委员会关于印发《xx市开展党员干部和公职人员涉酒违纪违法问题专项整治工作方案》的通知(市纪...
【其他范文】 日期:2022-11-08
-
班主任工作总结【2009年三年级语文教研组工作总结】
董晓玲本教研组共有6位语文教师,老中青结合,搭配合理,互帮互助,共同进步。我们坚持学校的有关教育教学计划,在学校教导处的领导下,严格执行学校的各项教育教学制度和要求...
【节日庆典】 日期:2019-08-02
-
理论中心组学习总体国家安全观发言材料9篇
理论中心组学习总体国家安全观发言材料9篇理论中心组学习总体国家安全观发言材料篇1(八)深入学习贯彻中央以及省的重要会议和文件精神深入学习贯彻年度内中央以
【发言稿】 日期:2022-08-04
-
军转座谈会交流发言4篇
军转座谈会交流发言4篇军转座谈会交流发言篇1大家好,我叫贺丽,2015届选调生,来自康定市委组织部,现在省委编办跟班学习。今天,非常荣幸向大家汇报我的学习收
【发言稿】 日期:2022-10-27
-
12岁生日小寿星发言4篇
12岁生日小寿星发言4篇12岁生日小寿星发言篇1各位来宾、各位朋友:大家好!今天,我们欢聚在这里,共同庆祝**十二周岁生日。首先,我代表**的父母以
【发言稿】 日期:2022-07-31
-
党内警告处分表态发言14篇
党内警告处分表态发言14篇党内警告处分表态发言篇1尊敬的各位领导、同事们:大家上午好!刚才会上宣布了党委关于我任职的决定,我首先衷心感谢党委的信任和
【发言稿】 日期:2022-09-13
-
党内警告处分党员讨论发言3篇
党内警告处分党员讨论发言3篇党内警告处分党员讨论发言篇1大家好!作为新时期的一名大学生,认真学习、深刻领会、全面贯彻省党代会精神,是当前和今后一个时期重
【发言稿】 日期:2022-08-07
-
【企业疫情风险控制方案】 2020企业复工疫情方案
企业疫情风险控制方案2020新冠病毒肺炎疫情防控工作总结汇报3篇 关于新型冠状病毒感染的肺炎疫
【演讲稿】 日期:2020-02-27
-
廉政大会总结发言稿7篇
廉政大会总结发言稿7篇廉政大会总结发言稿篇1各位领导,同志们:根据会议安排,我就党风廉政建设工作做表态发言,不妥之处,请批评指正。一、提高认识,切实
【发言稿】 日期:2022-10-30
-
被约谈的表态发言8篇
被约谈的表态发言8篇被约谈的表态发言篇1各位领导、各位党员大家好:这天我能站在鲜红的党旗下,
【发言稿】 日期:2022-12-24
-
破冰提能大讨论个人发言4篇
破冰提能大讨论个人发言4篇破冰提能大讨论个人发言篇1党史学习教育开展以来,我坚持读原著、学原文、悟原理。今天,根据会议安排,现在我就“学史明理”主题谈几点个
【发言稿】 日期:2022-10-09
-
巡察整改专题民主生活会总结发言8篇
巡察整改专题民主生活会总结发言8篇巡察整改专题民主生活会总结发言篇1按照区委统一部署和纪监委、巡察办关于召开党史学习教育专题组织生活会的工作安排,近期我紧贴
【发言稿】 日期:2022-10-12
-
2023年中国行政区划调整方案(设想优秀3篇
中国行政区划调整方案(设想优秀民政部第二次行政区划研讨会会议内容一、缩省的意义与原则1.意义1)利于减少中间层次中国行政区划层级之多为世界之最,既使管理成本
【周公解梦】 日期:2024-02-20
-
学习周永开先进事迹心得体会3篇
学习周永开先进事迹心得体会【一】通过学习周永开老先生先进事迹后,结合自己工作思考,感慨万千。同样作为
【格言】 日期:2021-04-10
-
2023年和儿媳妇在一起幸福的句子3篇
和儿媳妇在一起幸福的句子1、假如人生不曾相遇,我还是那个我,偶尔做做梦,然后,开始日复一日的奔波,淹没在这喧嚣的城市里。我不会了解,这个世界还有这样的一个你
【格言】 日期:2023-11-10
-
最满意的三项工作200字【最新党办公务员副主任提拔考察个人三年思想工作总结报告】
党办公务员个人三年工作总结近三年来,本人在组织、领导的关心指导和同事们的团结协作下,尽快完成主角的转
【格言】 日期:2021-02-26
-
XX老干局推进党建与业务深度融合发展工作情况调研报告:党建调研报告
XX老干局推进党建与业务深度融合 发展工作情况的调研报告 党建工作与业务工作融合发展始终是一个充满生
【成语大全】 日期:2020-08-28
-
中国共产党第三代中央领导集体的卓越贡献
中国共产党第三代中央领导集体的卓越贡献 --------------继往开来铸就辉煌 【摘要】改
【成语大全】 日期:2020-03-20
-
信息技术2.0能力点 [全国中小学教师信息技术应用能力提升工程试题题库及参考答案「精编」]
全国中小学教师信息技术应用能力提升工程试题题库及答案(复习资料)一、判断题题库(A为正确,B为错误)
【格言】 日期:2020-11-17
-
党建工作运行机制内容有哪些_构建基层党建工作运行机制探讨
党的基层组织是党在社会基层组织中的战斗堡垒,是党的全部工作和战斗力的基础。加强和改进县级以下各类党的
【经典阅读】 日期:2020-01-22
-
电大现代教育原理_最新国家开放大学电大《现代教育原理》形考任务2试题及答案
最新国家开放大学电大《现代教育原理》形考任务2试题及答案形考任务二一、多项选择题(共17道试题,共3
【成语大全】 日期:2020-07-20
-
集合推理_七,推理与集合
七推理与集合1 期中考试数学成绩出来了,三个好朋友分别考了88分,92分,95分。他们分别考了多少分
【名人名言】 日期:2020-12-18
-
关于三农工作重要论述心得体会3篇
关于三农工作重要论述心得体会3篇关于三农工作重要论述心得体会篇1习近平总书记指出:“建设现代化国家离不开农业农村现代化,要继续巩固脱贫攻坚成果,扎实推进乡村
【学习心得体会】 日期:2022-10-29
-
【福生庄隧道坍塌处理方案】 福生庄隧道在哪里
(呼和浩特铁路局大包电气化改造工程指挥部,内蒙古呼和浩特010050)摘要:文章介绍了福生庄隧道
【学习心得体会】 日期:2020-03-05
-
五个一百工程阅读心得体会13篇
五个一百工程阅读心得体会13篇五个一百工程阅读心得体会篇1凡益之道,与时偕行。在全国网络安全和信
【学习心得体会】 日期:2022-12-07
-
城管系统警示教育心得体会9篇
城管系统警示教育心得体会9篇城管系统警示教育心得体会篇1各党支部要召开多种形式的庆七一座谈会,组织广大党员进行座谈,回顾党的光辉历程,畅谈党的丰功伟绩,
【学习心得体会】 日期:2022-10-09
-
发展对象培训主要内容10篇
发展对象培训主要内容10篇发展对象培训主要内容篇1怀着无比激动的心情,我有幸参加了__新区区委党校20__年第四期(区级机关)党员发展对象培训班。这次的学习
【培训心得体会】 日期:2022-09-24
-
扶眉战役纪念馆心得体会11篇
扶眉战役纪念馆心得体会11篇扶眉战役纪念馆心得体会篇1有那么一段历史,低诉着血和泪的故事,慢慢地,随岁月老去;有那么一群人,放弃了闲逸的人生,辗转奔波中
【学习心得体会】 日期:2022-08-03
-
凝聚三种力量发展全过程人民民主心得体会12篇
凝聚三种力量发展全过程人民民主心得体会12篇凝聚三种力量发展全过程人民民主心得体会篇1新民主主义革命是指在帝国主义和无产阶级革命时代,殖民地半殖民地国家中的
【学习心得体会】 日期:2022-08-31
-
2022年全国检察长会议心得7篇
2022年全国检察长会议心得7篇2022年全国检察长会议心得篇1眼睛是心灵上的窗户,我们通过眼睛才能看到世间万物,才能看到眼前这美好的一切。拥有一双明亮的眼
【学习心得体会】 日期:2022-10-31
-
全面从严治党的心得体会800字7篇
全面从严治党的心得体会800字7篇全面从严治党的心得体会800字篇1中国特色社会主义是我们党领导
【学习心得体会】 日期:2022-12-14
-
教师全国两会精神学习专题研讨交流材料6篇
教师全国两会精神学习专题研讨交流材料6篇教师全国两会精神学习专题研讨交流材料篇1通过对两会精神深入系统的学习,作为新一代的青年人,更应该严格要求自己,贯彻落
【教师心得体会】 日期:2022-08-11
-
 2024年主题教育民主生活会批评与自我批评意见(38条)(范文推荐)
2024年主题教育民主生活会批评与自我批评意见(38条)(范文推荐)
2023年主题教育民主生活会六个方面个人检视、相互批评意见:1 理论学习系统性不强。学习习近平新时代中国特色社会主义思想不深不透,泛泛而学的时候多,深学细照的时候少,特...
【邓小平理论】 日期:2024-03-19
-
 2024年交流发言:强化思想理论武装,增强奋进力量(完整)
2024年交流发言:强化思想理论武装,增强奋进力量(完整)
习近平总书记指出:“一个民族要走在时代前列,就一刻不能没有理论思维,一刻不能没有思想指引。”党的十八大以来,伴随着新时代中国特色社会主义思想在实践中形成发展的历程...
【三个代表】 日期:2024-03-19
-
2024年度镇年度县乡人大代表述职评议活动总结
xx镇20xx年县乡人大代表述职评议活动总结为响应县级人大常委会关于开展县乡两级人大代表述职评议活动,进一步激发代表履职活力,加强代表与人民群众的联系,提高依法履职水平...
【马克思主义】 日期:2024-03-19
-
 “千万工程”经验学习体会(研讨材料)
“千万工程”经验学习体会(研讨材料)
“千万工程”是总书记在浙江工作时亲自谋划、亲自部署、亲自推动的一项重大决策,也是习近平新时代中国特色社会主义思想在之江大地的生动实践。20年来,“千万工程”先后经历...
【三个代表】 日期:2024-03-19
-
 2024年在市政协机关工作总结会议上讲话
2024年在市政协机关工作总结会议上讲话
同志们:刚才,XX同志对市政协机关20XX年工作进行了很好的总结,很精炼,很到位,可以感受到去年机关工作确实可圈可点。XX同志宣读了表彰决定,机关优秀人员代表、先进集体代...
【邓小平理论】 日期:2024-03-18
-
 在全区防汛防涝动员暨河长制工作推进会上讲话提纲【完整版】
在全区防汛防涝动员暨河长制工作推进会上讲话提纲【完整版】
区长,各位领导,同志们:汛期已经来临,我区城区防涝工作面临强大考验,形势不容乐观。年初,区城区防涝排渍指挥部已经召开专题调度会,修订完善应急预案,建立网格化管理机...
【马克思主义】 日期:2024-03-18
-
 2024年镇作风整治工作实施方案(完整文档)
2024年镇作风整治工作实施方案(完整文档)
XX镇作风整治工作实施方案为深入贯彻落实党的二十大精神及省市区委深化作风建设的最新要求,突出重点推进干部效能提升,坚持不懈推动作风整治工作纵深发展,根据《关于印发《2...
【毛泽东思想】 日期:2024-03-18
-
2024市优化法治化营商环境规范涉企行政执法实施方案【优秀范文】
xx市优化法治化营商环境规范涉企行政执法实施方案为持续优化法治化营商环境,激发市场主体活力和社会创造力,规范行政执法行为,创新行政执法方式,提升行政执法质效,着力解...
【毛泽东思想】 日期:2024-03-18
-
 2024年度关于开展新一轮思想状况摸底排查工作通知(完整)
2024年度关于开展新一轮思想状况摸底排查工作通知(完整)
关于开展新一轮思想状况摸底排查工作的通知为深入贯彻落实关于各地开展干部职工思想状况大摸底大排查情况上的批示要求和改革教育第二次调度会议精神,有针对性做好队伍教育管...
【三个代表】 日期:2024-03-18
-
2024年公路养护中心主任典型事迹材料(完整文档)
“中心的工作就是心中的事业”——公路养护中心主任典型事迹材料**,男,1976年6月出生,1993年参加工作,2000年4月调入**区交通运输局工作,大学本科学历,中共党员,现任**...
【马克思主义】 日期:2024-03-17